Master Bioinfo Projets et Stages
Désolé, cette activité n’est pas visible actuellement
Résumé de section
-
-
Les étudiants doiventMarquer comme terminé
-
Les étudiants doiventMarquer comme terminé
-
Les étudiants doiventMarquer comme terminé
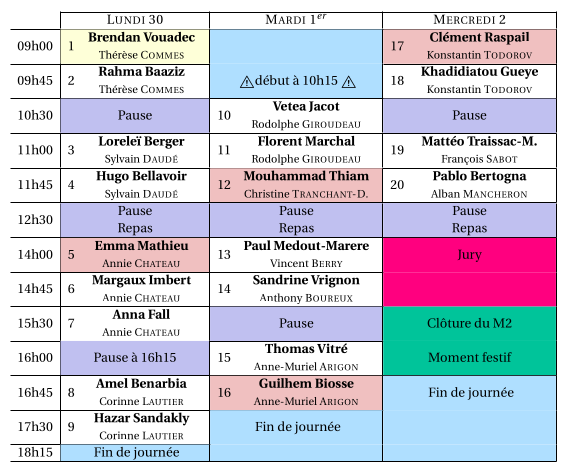
-
Annonces Forum
-
Les étudiants doiventMarquer comme terminé
-
Offres de stage 2024-25 Dossier
-
Offres de stage spéciales M1 2024-25 Dossier
-
Affectation tuteurs et tutrices M1 et M2 AS Fichier
-
Affectation tuteurs et tutrices M2 Fichier
-
-
-
Les étudiants doiventMarquer comme terminé

-
Les étudiants doiventMarquer comme terminé
-