Le système immunitaire
| Site: | Moodle UM - Plateforme pédagogique de l'Université de Montpellier |
| Cours: | L'histologie de A à Z |
| Livre: | Le système immunitaire |
| Imprimé par: | Visiteur anonyme |
| Date: | lundi 22 juillet 2024, 18:05 |
Description
1. INTRODUCTION
Le système immunitaire est un système très complexe qui nous protège des agressions extérieures qui peuvent être biologiques (virus, bactéries champignons, parasites), chimiques (ex : venins, toxines) ou physiques (traumatisme, brûlure, gelure, coup de soleil, …) et des agressions intérieures (cellules infectées ou cellules tumorales). Le système immunitaire est formé d’organes lymphoïdes (Figure 1), de cellules de l’immunité (Figure 2) et de protéines solubles (Figure 7). Les organes lymphoïdes sont répartis dans l’ensemble de l’organisme et ils sont physiquement reliés entre eux par les vaisseaux lymphatiques dans lesquels transitent les cellules de l’immunité et les protéines solubles à l’origine d’un dialogue permanent entre les organes lymphoïdes. Pour être efficace le système immunitaire est finement régulé par les cellules de l’immunité et les protéines solubles. Ceci afin d’éviter une diminution de la surveillance immunitaire ou une réaction trop excessive du système immunitaire conduisant à des situations pathologiques telles que les maladies auto-immunes, l’inflammation chronique ou l’allergie.
L’immunité de l’organisme est assurée de manière coordonnée et intégrée par l’Ante-immunité, l’immunité innée et l’immunité adaptative.
- Ante-immunité (= mécanisme de protection superficielle) : Barrière physique et chimique assurée par les cellules épithéliales de la peau et des différentes muqueuses de l’organisme (Figure 3). Cette première protection fait intervenir essentiellement les cellules épithéliales. Outre les cellules épithéliales, les cellules endothéliales et les plaquettes jouent un rôle relais entre les mécanismes de protection superficielle et l’immunité innée.
- Immunité innée : Contribue à l’élimination des agents pathogènes, ayant pénétré à l’intérieur des tissus. Cette élimination se fait grâce aux cellules de l’immunité innée (Figure 5) : les polynucléaires, (Basophile, Eosinophile et Neutrophile), les phagocytes mononucléés (macrophages et cellules dendritiques), les mastocytes et les cellules lymphoïdes non conventionnelles ainsi qu’aux protéines solubles (cytokines et protéine du complément)
- Immunité adaptative : Intervient en appui de l’immunité innée pour éliminer les corps étrangers persistants. Les principaux acteurs cellulaires sont les lymphocytes qui interagissent avec les cellules de l’immunité innée. Parmi les acteurs protéiques solubles, on distingue les cytokines et les anticorps.
Lors d’une agression extérieure (ex infection par une bactérie) les trois composantes du système immunitaire vont s’articuler autour d’une réaction biologique, la réaction inflammatoire. Dans cette réaction, la composante « système immunitaire innée » est majoritaire et elle fonctionne de manière intégrée avec la composante « Ante-immunité » et la composante « système immunitaire adaptatif ».
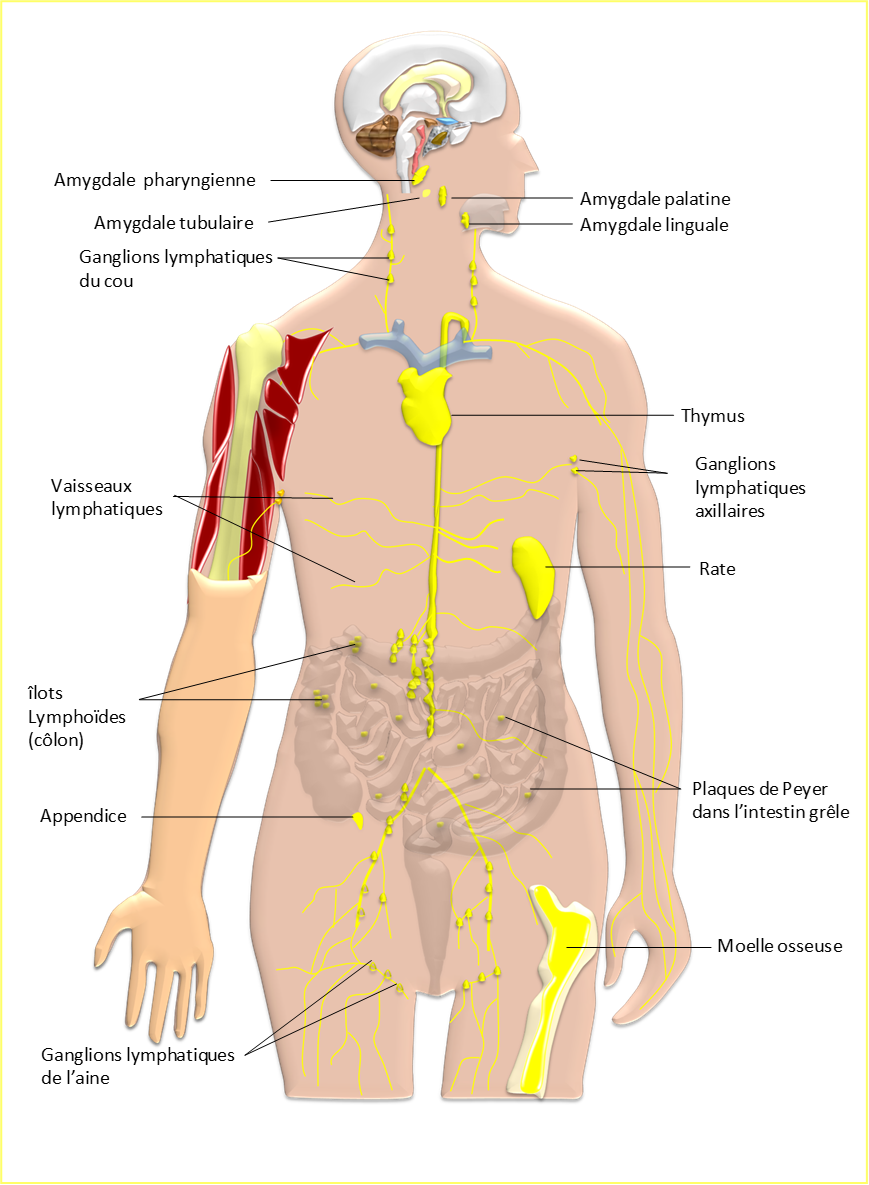
Figure
1 : les organes lymphoïdes du système immunitaire
https://online.fliphtml5.com/dhphd/sbvd/index.html
Accéder à la version papier
Bibliothèque de la faculté de médecine Montpellier et Nîmes
1.1. Ante - immunité
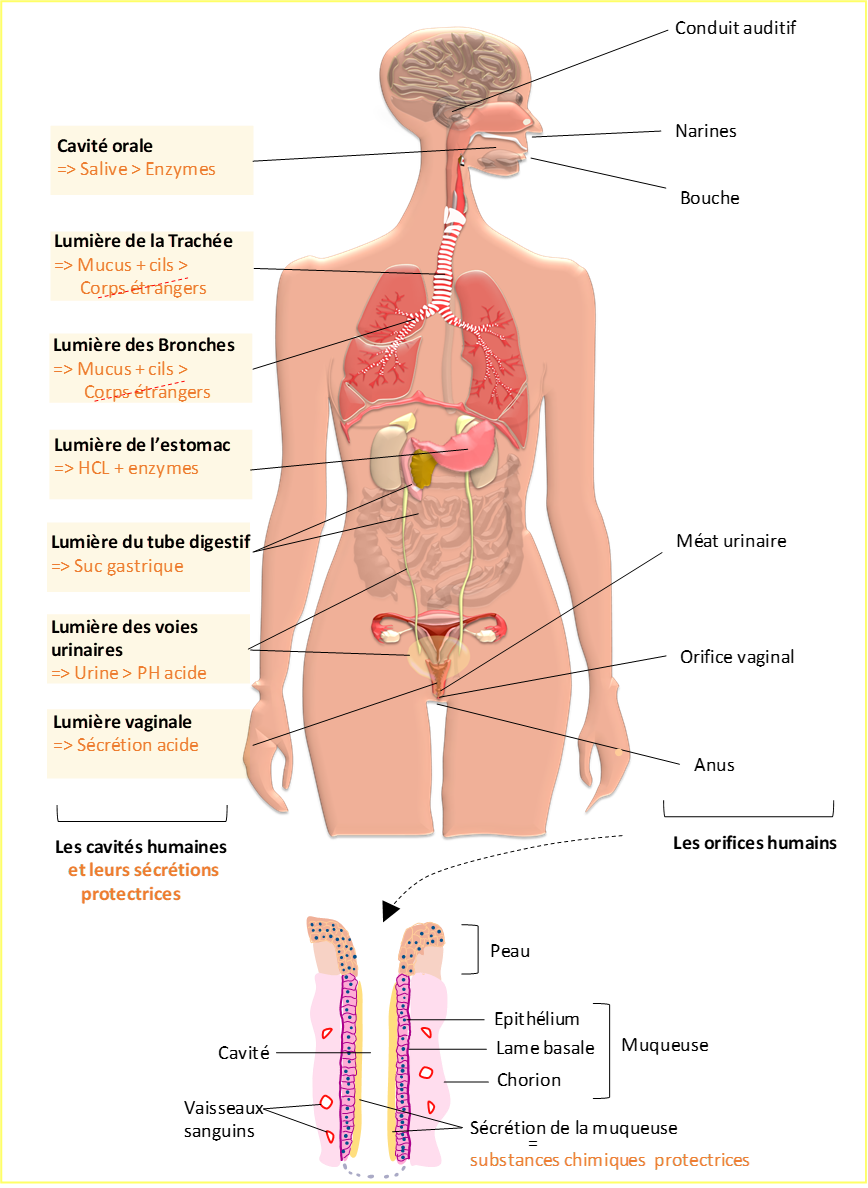
Pour vivre en symbiose avec le monde extérieur, notre organisme est entièrement recouvert d’un revêtement cutané : la peau. Par endroits, la peau est interrompue par la présence d’orifices qui permettent à l’organisme de communiquer avec l’environnement extérieur (Figure 3). Ces orifices sont le point d’entrée de cavités (espaces vides) qui pénètrent à l’intérieur du corps humain. Ces cavités sont tapissées par une muqueuse formée successivement de l’extérieur vers l’intérieur par un épithélium, une lame basale et un tissu conjonctif richement vascularisé appelé le chorion. La principale cavité est formée par le tube digestif qui s’étend de la bouche à l’anus. La deuxième cavité assure la respiration et débute des narines et de la bouche jusqu’aux alvéoles des poumons. Chez la femme une importante cavité prend naissance au niveau du vagin et se poursuit jusqu’aux ovaires. L’oreille est formée d’une cavité qui nous permet de percevoir les sons. Enfin l’appareil urinaire est parcouru par un réseau de cavités plus ou moins larges : la vessie, le rein et les réseaux tubulaires permettant la formation et l’élimination des urines. Chaque cavité présente une muqueuse spécifique qui s’adapte aux fonctions des organes qu’elle recouvre.
L’épithélium de la peau et des muqueuses constitue une barrière physique contre les micro-organismes et, grâce à leurs sécrétions diverses, une barrière chimique empêchant les micro-organismes de prospérer à leur surface et de les traverser.
Parmi les différentes sécrétions il existe :
- les sécrétions de la peau (sueur et sébum) rendent la surface de l’épithélium acide permettant d’inhiber la prolifération des bactéries
- les glandes salivaires produisent la salive contenant des enzymes, les lysosomes, qui détruisent les bactéries
- les muqueuses gastriques de l’estomac sécrètent une solution, appelée suc gastrique, qui détruit la plupart des micro-organismes. Elle est composée d’acide (l’acide chlorhydrique (HCL)) et de protéases (enzymes qui dégradent les protéines)
- le mucus produit par les muqueuses respiratoires et notamment par les cellules caliciformes et les acini muqueux. Le mucus présent à la surface de l’épithélium respiratoire piège les micro-organismes et les particules inférieures à 7µm. Les cils des cellules ciliées, par un mouvement coordonné, transportent les micro-organismes ainsi piégés dans le mucus jusqu’au pharynx où ils seront soit expectorés soit avalés pour être acheminés jusqu’à l’estomac où ils seront éliminés
- les sécrétions vaginales acides contribuent à éliminer de nombreux micro-organismes pathogènes qui tentent de s’implanter dans l’appareil génital féminin
- l’urine présente un PH acide qui inhibe la prolifération bactérienne
Outre leurs fonctions de barrières physiques et chimiques, les cellules épithéliales des épithéliums de la peau ou des muqueuses sont les premières cellules résidentes ayant la capacité de détecter les signaux de dangers (DAMP : Danger Associated Molecular Pattern), les signaux pathogéniques (MAMP : Microbe associated Molecular patterns ; (Figure 4) et de sécréter en conséquence des cytokines; ces dernières activent les cellules résidentes du système immunitaire inné (cellules qui résident dans le tissu : Macrophages, …). Des fonctions similaires ont été également décrites à partir des cellules endothéliales et des plaquettes.
1.2. Le système immunitaire inné
1.2.1 Introduction
Le système immunitaire inné reconnaît, grâce à ces différents récepteurs PPRs (Pattern Recognition Receptors; Figure 4), des structures moléculaires spécifiques portées par des agents infectieux, regroupés sous l’entité MAMP (Microbe Associated Molecular Patterns) ou produites par des tissus lésés regroupés sous l’appellation DAMP (Damage Associated Molecular Patterns). L’activation du système immunitaire inné par ces structures moléculaires se caractérise par une réponse locale, rapide et peu spécifique qui lui permet ainsi de combattre un large panel d’agents pathogènes. Dans le système immunitaire inné il n’y a pas de processus d’apprentissage : l’amplitude de la réponse est identique à chaque nouvelle rencontre de l’agent pathogène.
Le système immunitaire inné est constitué de cellules et de messagers solubles. Il contribue pour une large part à l’initiation de la réaction inflammatoire aiguë qui constitue la toute première réponse se déroulant dans le tissu, suite à la détection de signaux de dangers ou pathogéniques par les cellules résidentes également appelées cellules sentinelles.
1.2.2 Les acteurs cellulaires
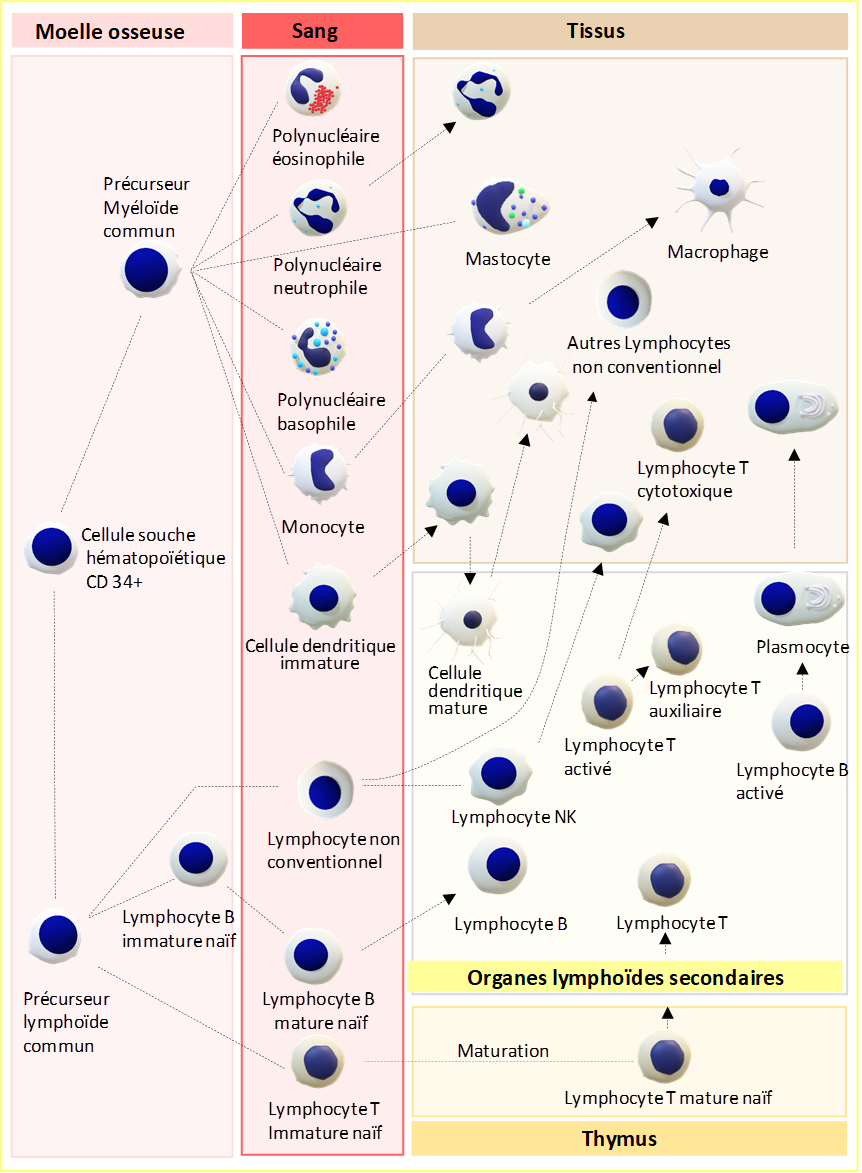
Toutes les cellules du système immunitaire inné sont issues de la moelle osseuse et dérivent soit d’un précurseur myéloïde commun soit d’un précurseur lymphoïde commun, provenant tous deux de la différenciation des cellules souches hématopoïétiques CD34+ (Figure 2). La majorité provient de la lignée myéloïde (Figures 2 et 5) :
- les polynucléaires : neutrophiles, basophiles et éosinophiles
- les phagocytes mononucléés : les monocytes/macrophages et les cellules dendritiques
- les mastocytes
D’autres sont d’origines lymphoïdes avec la particularité d’exprimer un récepteur TCR (Figures 13, 14 et 15) non conventionnel. Ces lymphocytes sont regroupés sous l’appellation : lymphocytes T non conventionnels.
Toutes ces cellules migrent de la moelle osseuse vers le sang périphérique. Certaines d’entre elles quitteront les vaisseaux sanguins pour résider dans les tissus, c’est par exemple le cas des cellules dendritiques, des mastocytes et des monocytes. Ces derniers se transforment en macrophages dans les tissus.

Figure
4 : Structure moléculaire détectée
par le système immunitaire : MAMP
et DAMP

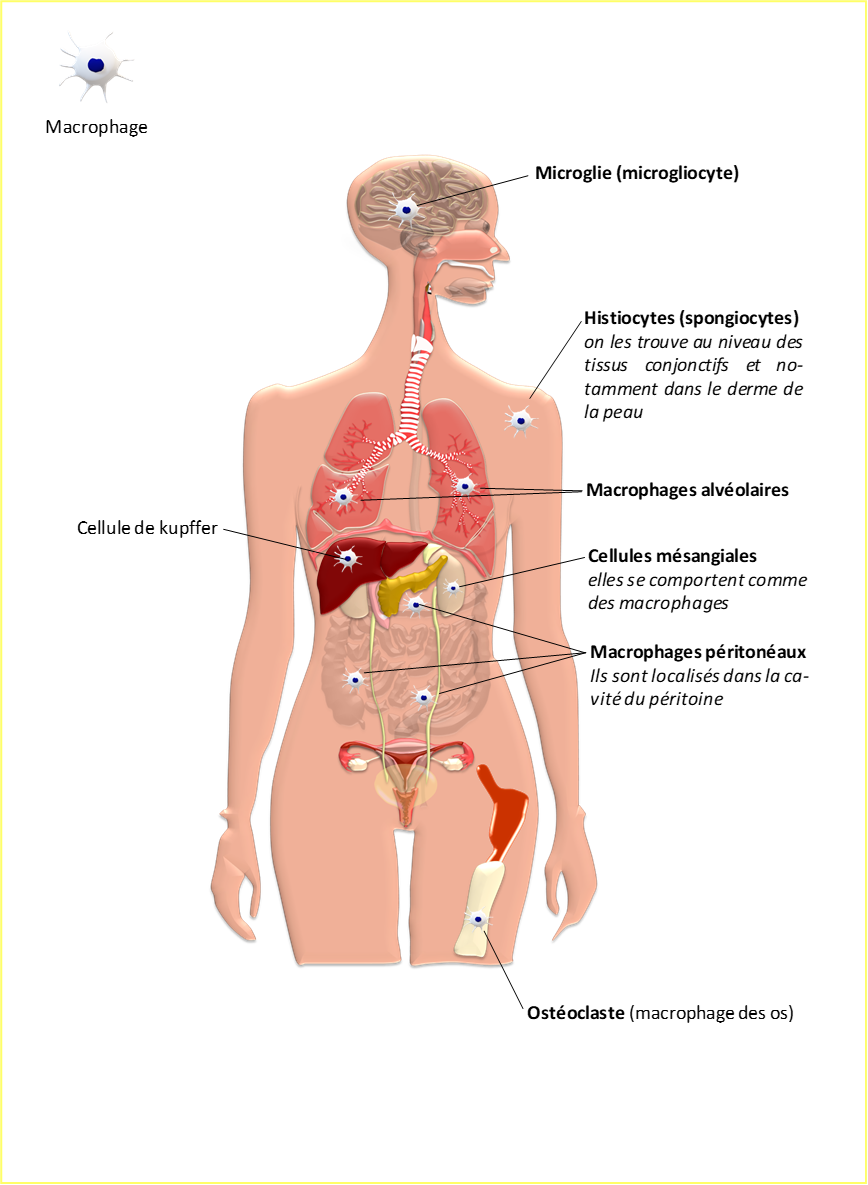
Figure 6 : Nomenclature associée aux macrophages en fonction des tissus colonisés

Figure 7 : Cytokines du système immunitaire inné
Lors de réactions inflammatoires aiguës ou chroniques, les polynucléaires et ou les monocytes colonisent les tissus lésés.
Chaque cellule du système immunitaire inné possède des propriétés et des fonctions bien définies (Figure 5).
1.2.3 Les acteurs protéiques
1.2.3.1 Les cytokines de l’inflammation
Les cellules du système immunitaire inné interagissent entre elles et avec le milieu environnant au travers de cytokines (du grec « cyto », cellule et « kinos », mouvement) qu’elles sécrètent (Figure 7). Contrairement aux hormones, les cytokines exercent leur fonction majoritairement dans un environnement local. Elles sont sécrétées suite à un stimulus externe (ex : détection d’un corps étrangers) puis elles se fixent à leur cellule cible par l’intermédiaire d’un récepteur membranaire. Elles permettent ainsi de réguler la réponse immunitaire. En fonction de leur rôle, on distingue plusieurs types de cytokines (Figure 7).
1.2.3.2 Les protéines du complément
Le complément est un élément central de l’immunité inné. Il constitue la première ligne de défense contre les agents pathogènes. Le complément agit dans l’ensemble de l’organisme au niveau des tissus, des vaisseaux sanguins et lymphatiques et à l’intérieur des cellules. Il peut être déclenchée par 3 voies différentes, la voie classique, la voie des lectines et la voie alternative (Fiche 1 et ses commentaires, p152). Le complément est formé d’une cinquantaine de protéines plasmatiques qui sont soit en phase liquide soit associées aux membranaires des cellules cibles. Les protéines du complément sont impliquées dans la détection des agents pathogènes et leur destruction (Fiches 2 et 3 et ses commentaires, p153) soit directement (complexe d’attaque membranaire) ou indirectement (induction de l’opsonisation, de l’inflammation et de l’activation du système immunitaire adaptatif).
1.2.4 Le système immunitaire inné dans l’inflammation aigüe
Le terme général d’inflammation a été affiné et remplacé par l’inflammation aiguë qui correspond à une inflammation contrôlée, adaptée, permettant à l’organisme de combattre les corps étrangers et réparer les tissus ; versus l’inflammation chronique qui est une inflammation incontrôlée et agressive conduisant à des situations pathologiques.
La réaction inflammatoire aiguë comporte 3 phases principales : la phase d’initiation, la phase d’amplification et la phase de réparation.
Dans le chapitre suivant nous traiterons ces différentes phases dans le contexte d’une infection bactérienne se déroulant au niveau d’une muqueuse.
1.2.5 La phase d’initiation de l’inflammation aigüe
Elle débute (Figure 8) par l’apparition de signaux pathogéniques (MAMP) et de signaux de dangers (DAMP) captés par les cellules résidentes que sont les macrophages, les cellules dendritiques, les mastocytes et / ou par le système du complément (Fiches 4 et ses commentaires : Pour en savoir +, p153). Ces signaux peuvent également être détectés par les cellules épithéliales, endothéliales et les plaquettes. Il en résulte une activation des cellules résidentes qui se traduit par :- la sécrétion de cytokines responsables
du recrutement des cellules du système immunitaire inné (neutrophiles, puis
macrophages)
- la vasodilatation des vaisseaux sanguins
- l’activation du système du complément
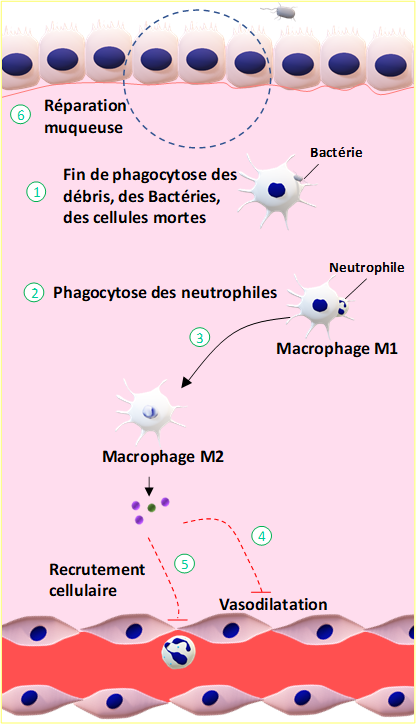
Figure 8 : L’inflammation aigüe : la phase d’initiation. Des lésions tissulaires créées par les bactéries au niveau de la muqueuse respiratoire ①, génèrent des signaux activateurs ②. Ces signaux activent les cellules résidentes (macrophages, mastocytes, cellules dendritiques ③ qui produisent et sécrètent des cytokines ④, induisant le recrutement cellulaire ⑤ et la vasodilatation des vaisseaux sanguins ⑥.
1.2.6 La phase d’amplification de l’inflammation aigüe
Les neutrophiles sont les premières cellules du système immunitaire inné à être recrutées au niveau du tissu lésé pour détruire les corps étrangers, ici en l’occurrence les bactéries (Fiches 5 et ses commentaires : Pour en savoir +, p153). Sont ensuite recrutés les cellules dendritiques et les monocytes circulants qui se transforment en macrophages lors de leur passage dans les tissus lésés. Dès leur arrivée au sein des tissus lésés, les neutrophiles, les cellules dendritiques et les macrophages sont activés soit par les cytokines produites par les cellules résidentes soit par interaction direct avec les bactéries soit par les protéines du système du complément. Les neutrophiles, macrophages et cellules dendritiques procèdent ensuite à l’élimination des bactéries par différents mécanismes : l’opsonisation, la phagocytose, la dégranulation, l’explosion oxydative, et la nétose (Fiches 5 et ses commentaires, p153). Le système du complément contribue à la mise en place de ces différents mécanismes et, de plus, il est directement responsable de la formation du complexe d’attaque (Fiches 2 et ses commentaires, p153) qui constitue un mécanisme supplémentaire pour la destruction des corps étrangers.
Dans le même temps, les cellules dendritiques immatures ayant capturés les corps étrangers migrent en direction des ganglions lymphatiques et présentent les déterminants antigéniques issus des antigènes capturés aux lymphocytes T et B.
Si les corps étrangers sont éliminés, la production et la sécrétion de cytokines par les cellules de l’immunité innée cessent, marquant l’arrêt de la réponse inflammatoire et le début de la phase de réparation tissulaire. Ce moment clef de la réaction inflammatoire correspond à la résolution de l’inflammation.
1.2.7 La phase de réparation tissulaire de l’inflammation aigüe
Après élimination des agents pathogènes (Figure 9), des débris cellulaires et des cellules mortes par les cellules phagocytaires, les neutrophiles rentrent en apoptose et sont phagocytés à leur tour par les macrophages, processus appelé efférocytose (Fiches 6 et ses commentaires, p153). A ce stade, la réaction inflammatoire aiguë entre dans une phase décroissante qui se traduit par un arrêt de la production de cytokines pro-inflammatoires au profit de cytokines anti-inflammatoires. Ce changement induit l’arrêt de recrutement des cellules immunitaires, l’inhibition des cellules du système inné présentes sur le site lésé et l’initiation des mécanismes de réparation tissulaire.La réparation des tissus est assurée par différents types cellulaires en fonction des dégâts. En cas d’une lésion mineure, les cellules endothéliales produisent la matrice extracellulaire permettant la réparation des tissus. Pour des lésions plus graves, les fibroblastes produisent la matrice extracellulaire et les macrophages participent à l’angiogenèse (création de nouveaux vaisseaux sanguins).
1.3. Le système immunitaire adaptatif
Le système immunitaire adaptatif est doté
d’une mémoire qui lui assure une capacité d’apprentissage. Dès qu’il rencontre un
agent pathogène, le système immunitaire adaptif enregistre ce premier contact
pour adapter au mieux, lors d’une deuxième rencontre, sa réponse en termes
d’intensité, de spécificité, de rapidité et de puissance, dans le but
d’éliminer le plus efficacement possible l’agent en question. Le système
immunitaire inné participe à l’activation du système adaptatif qui repose sur
deux acteurs principaux : les lymphocytes B à l’origine de la réponse
humorale et les lymphocytes T pour la réponse cellulaire.
En réalité, les deux systèmes immunitaires inné et adaptatif sont étroitement imbriqués. Ainsi ils se régulent et se contrôlent mutuellement. Le système immunitaire inné régule le système immunitaire adaptatif via ses cellules présentatrices d’antigènes (macrophages et cellules dendritiques). A l’inverse, le système adaptatif contrôle le système inné lors de la réaction humorale par exemple : les lymphocytes B se différencient en plasmocytes qui produisent des anticorps. Ceux-ci peuvent recouvrir des agents pathogènes comme les bactéries favorisant leur phagocytose par les cellules phagocytaires du système immunitaire inné.
Le système immunitaire adaptatif est formé par deux acteurs majeurs : les cellules et les protéines.
1.3.1 Les acteurs cellulaires
Les cellules du système immunitaire adaptatif et les cellules du système immunitaire inné sont issues de la moelle osseuse. Les cellules souches hématopoïétiques CD34+ de la moelle osseuse se différencient en un précurseur lymphoïde commun qui génère à son tour deux types de lymphocytes, les précurseurs des lymphocytes B et T (Figure 10). Les précurseurs des lymphocytes B poursuivent leur maturation dans la moelle osseuse jusqu’au stade de lymphocyte B immature. En revanche, les précurseurs des lymphocytes T quittent la moelle osseuse via les vaisseaux sanguins et vont poursuivre leur maturation dans le thymus (Figures 10 et 25 et chapitre 2.2.2) pour générer deux sous-populations de lymphocytes T (LT), les lymphocytes T CD4+ naïfs et les lymphocytes T CD8+ naïfs.
Les lymphocytes B immatures quittent la moelle osseuse à leur tour et poursuivent leur maturation dans le sang périphérique puis dans les organes lymphoïdes secondaires (Ganglions lymphatiques et Rate) tout comme les lymphocytes T CD4+ naïfs et T CD8+ naïfs.
C’est dans les organes lymphoïdes secondaires, présentant une architecture très structurée, que les lymphocytes T CD4+ et T CD8+ naïfs et les lymphocytes B matures peuvent rencontrer leur antigène (Figure 11) grâce respectivement à leurs récepteurs BCR (Figure 12) et TCR (Figure 13). Les lymphocytes T CD4+ et T CD8+ naïfs se distinguent respectivement par l’ajout des co-récepteurs CD4 (Figure 14) et CD8 (Figure 15) au complexe TCR.

Figure 10 : Les cellules du système immunitaire adaptatif
Alors que les lymphocytes B reconnaissent les antigènes
dans leur forme native, les lymphocytes T les reconnaissent sous forme de
peptides présentés par le complexe majeur d’histocompatibilité (CMH) de classe
I (Fiche 7) pour
les LT CD8+ et de classe II pour les CD4+ (Fiche 8). Les lymphocytes T CD4+ naïfs
peuvent se différencier une fois activés en divers types de lymphocytes Th en
fonction des cytokines polarisantes ayant conduit à leur activation (Figure 10). Plusieurs types de
Lymphocytes Th ont été répertoriés dans la littérature en fonction des
cytokines produites et sécrétées par les lymphocytes Th.
Les principaux sont les suivants : les lymphocytes Th1, Th2, Th17 et les
Treg. D’autres lymphocytes Th comme les lymphocytes TFh, Th9, Th22, Th3 et Tr1
sont également proposés ; mais à ce jour, il n’a pas été établi avec
certitude s’ils sont distincts ou s’ils proviennent de
sous-types de Th1, Th2 ou Th17. Les
lymphocytes T CD8+ naïfs peuvent, quant à eux, s’activer et se différencier en
lymphocytes T CD8+ cytotoxiques (CTL). les lymphocytes B activés se différencient en lymphocytes
B mémoire et plasmocytes.


Figure 12: Le récepteur BCR des LB
1.3.2 Les acteurs protéiques
1.3.2.1 Les cytokines
Les cytokines du système adaptatif sont issues :
- des lymphocytes T CD4+ naïfs : ces derniers se différencient en lymphocytes Th spécifiques, chacun produisant et sécrétant une combinaison de cytokines données (Figure 10). Parmi ces cytokines, on distingue, des interleukines (IL-4, IL-21, IL-5, IL-13, IL-17, IL-22, …), l’interféron gamma (IFNƴ) et le facteur de croissance tumoral (TGFβ).
- des lymphocytes T CD8+ naïfs qui, une fois activés, se différencient en LT CD8+ cytotoxiques capables de produire et sécréter également de l’IFNƴ.
Figure 13 : TCR et complexe TCR

Figure 14 : Récepteur TCR : LT CD4+
1.3.2.2 Les enzymes
Les LT CD8+ cytotoxiques sécrètent également des perforines (enzymes qui perforent la membrane cytoplasmique de la cellule cible) et des granzymes A et B qui activent l’apoptose des cellules cibles.
1.1.1.2 Les anticorps
Les lymphocytes B activés se différencient en plasmocytes qui peuvent sécréter 5 classes principales d’immunoglobuline communément appelé anticorps : IgG, IgA, IgM, IgD et IgE. Sous forme soluble, l’immunoglobuline exerce sa fonction d’anticorps et, lorsqu’elle est ancrée à la membrane, contribue à la formation du récepteur BCR (Figure 12).
1.3.3 Le système immunitaire adaptatif dans l’inflammation aigüe
Si la réaction inflammatoire persiste, c’est-à-dire si les agents pathogènes ne sont pas éliminés (ici les bactéries : chapitre 1.2.5) , la phase de réparation tissulaire n’a pas lieu et le système immunitaire adaptatif prend part à la réaction inflammatoire. Les cellules dendritiques qui reconnaissent l’agent pathogène responsable de l’inflammation s’active et se transforme en cellules présentatrices d’antigènes (CPA). Elles expriment alors le récepteur CCR7 qui leur permet de pénétrer à l’intérieur des vaisseaux sanguins et de migrer en direction des organes lymphoïdes secondaires dans les zones riches en lymphocyte T. Les CPA qui ont capturés l’antigène soit par phagocytose, endocytose ou pinocytose internalise l’antigène, le digère au niveau des endosomes formant un peptide antigénique qui s’associe aux CMH II et qui est présenté aux Lymphocytes T CD4+ naïfs (Fiche 8). Si l’affinité est suffisante les lymphocytes T CD4+ naïfs s’activent et se différencient en lymphocytes T auxiliaire (Th). Les lymphocytes Th sont nécessaires à l’activation des lymphocytes B. L’activation des lymphocytes B et leur différentiation en plasmocyte (Figure 38) nécessite également une interaction avec son antigène (via les CPA ou sous forme soluble ou d’immuns complexes). Les plasmocytes sécrètent des anticorps qui diffusent notamment vers le tissu lésé et participe à l’élimination des bactéries en interaction avec le complément et les macrophages.
Figure 15 : Récepteur TCR : LT CD8+
1.4. Les organes du système immunitaire
Les organes du système immunitaire sont répartis dans l’ensemble de l’organisme et sont reliés entre eux via les réseaux de vaisseaux sanguins et lymphatiques (Figure 1). Les cellules des systèmes immunitaires adaptatif et inné empruntent ces réseaux et assurent ainsi la communication entre ses différents organes. Les organes du système immunitaire sont répartis en deux groupes : les organes lymphoïdes primaires et les organes lymphoïdes secondaires. Parmi les organes lymphoïdes primaires on distingue :
- la moelle osseuse essentiellement localisée dans les os courts et plats comme ceux du crâne, de la colonne vertébrale, du sternum, du bassin ou encore des côtes. C’est dans la moelle osseuse que les cellules souches hématopoïétiques CD34+ s’autorenouvellent et peuvent générer l’ensemble des cellules du système immunitaire (Figure 2). La moelle osseuse est aussi le lieu de maturation des lymphocytes B où ils acquièrent un récepteur BCR.
- le thymus situé dans le médiastin antérieur. Les lymphocytes T précurseurs en provenance de la moelle osseuse pénètrent dans le thymus où ils acquièrent un récepteur de surface TCR, puis les récepteurs CD4 ou CD8 pour former les lymphocytes T CD4+ naïfs ou les lymphocytes T CD8+ naïfs. Les lymphocytes T et B naïfs quittent les organes lymphoïdes primaires et circulent dans les vaisseaux sanguins et lymphatiques en direction des organes lymphoïdes secondaires où ils pourront rencontrer leur antigène correspondant pour finir leur maturation. Les organes lymphoïdes secondaires sont plus nombreux et recouvrent l’ensemble de l’organisme :
- les ganglions lymphatiques sont situés à la jonction des faisceaux lymphatiques qui drainent les tissus de l’ensemble de l’organisme. De petite taille (1-15 mm de diamètre), chaque individu en possède entre 500 et 1000. Les ganglions lymphatiques constituent le lieu préférentiel de rencontres entre les lymphocytes B et T matures naïfs, les antigènes et les cellules CPA de la lymphe circulante. Ils permettent ainsi l’activation des lymphocytes matures naïfs, sous condition que l’affinité entre l’antigène et le récepteur des lymphocytes soit suffisante.
- la rate est localisée dans le quadrant supérieur gauche de l'abdomen, en dessous de la cage thoracique. La rate permet aux lymphocytes B et T matures naïfs d’interagir avec les antigènes présents dans le sang circulant et ainsi les activer si affinité.
- le tissu lymphoïde associé aux muqueuses (MALT, pour Mucosal Asso-ciated Lymphoïd Tissue) est situé princi-palement dans les muqueuses respira-toires (amygdales et végétations adénoï-des de l’oropharynx), gastro-intestinales (îlots lymphoïdes et plaques de Meyer) et urogénitales. Ces structures lymphoïdes sont formées par des lymphocytes B et T matures et des plasmocytes qui peuvent interagir avec les antigènes ayant pénétré les muqueuses.2. PARTIE II : LES ORGANES LYMPHOÏDES PRIMAIRES
Accéder gratuitement à l'ensemble de l'ouvrage
https://online.fliphtml5.com/dhphd/sbvd/index.html
Accéder à la version papier
Le système immunitaire : histologie
3. PARTIE III : LES ORGANES LYMPHOÏDES SECONDAIRES
Accéder gratuitement à l'ensemble de l'ouvrage
https://online.fliphtml5.com/dhphd/sbvd/index.html
4. PARTIE IV : RESUME : LES ORGANES LYMPHOÏDES

Accéder gratuitement à l'ensemble de l'ouvrage
https://online.fliphtml5.com/dhphd/sbvd/index.html
5. PARTIE V : ANNEXE
Accéder gratuitement à l'ensemble de l'ouvrage
https://online.fliphtml5.com/dhphd/sbvd/index.html